Twins Separated At Birth: The Story of Asymmetric Cell Division
You’ve probably heard of mitosis and meiosis but what if I told you that a specific type of cell division has been hidden from you all this time? Cells DON’T always divide into two identical cells and it’s the reason why we’re still alive.

Let’s first imagine a crazy scenario. We’re usually taught that mitosis or cell division is when a mother or parent cell divides into two identical daughter cells, right?
But [then how come] instead of being a hundred or so pounds of cells in a bucket, we’re functioning human beings?
– Dr. Yukiko Yamashita, Associate Professor at the University of Michigan
It’s because that’s only a part of the story! What I just described–one cell dividing into two identical ones–is called symmetric cell division. But there’s another mechanism called asymmetric cell division or ACD where both cells don’t have the same cell fate! Before we fully understand why that is, we need to cover some basics.
The Basics
The Cell Family
First off, say hello to the cell family! We have the mother cell or parent cell and the two daughter/sister cells.
The mother cell is the initial cell where everything starts. Then from there, the mother cell divides into two new cells which are called daughter cells with respect to the mother cell and sister cells with respect to each other. That’s easy enough to understand!

The Cell Cycle: A Focus On Mitosis
The cell cycle is the process through which we get daughter cells from the mother cells or just the process which helps us create more cells so we can grow and repair damaged tissues.
There are two major stages of the cell cycle that have their own substages. And the two stages are interphase and mitosis.
Interphase is where the cell spends most of its life and it’s when DNA is duplicated, when extra proteins needed for division are created and more. Before we get to the more exciting part and the one we’ll be focusing on, mitosis, we need to double click on what DNA is. DNA is like the recipe of life but since we have a lot of DNA, we need to organize it somehow. Just like how text is organized into books and books into bookshelves, chromosomes are like the books of our DNA and the nucleus is the bookshelf. Chromosomes make it much easier to duplicate DNA and to move it around as we’ll see soon!

Just a quick note on something that might seem confusing as you continue studying the cell cycle: there are 46 chromosomes both before and after duplication. Huh? Well, looking at a drawing of chromosomes below, you can see that they have this centre, called the centromere. And we count chromosomes by the number of centromeres. Each of those little arms is a sister chromatid. So after duplication, we still have 46 chromosomes but 92 sister chromatids.

With that out of the way, it’s time to go back to the cell cycle! It’s important to note that most the time is spent in interphase but mitosis is where the fun begins! Zooming in further, we can see there are 4 substages in mitosis: prophase, metaphase, anaphase, and telophase. The acronym most people use to remember this is PMAT but here’s a nice mnemonic device you can use too:
P eter
M arches
A nd
T umbles
One interesting way to think about mitosis is like training an army. (Quick note that this is just an analogy and not an accurate representation of what it takes to be in the military!) In early prophase not too much is happening but the chromosomes begin condensing or thickening so that in later stages, they don’t break or get tangled while being moved around. Think of it like when the soliders just begin their training.
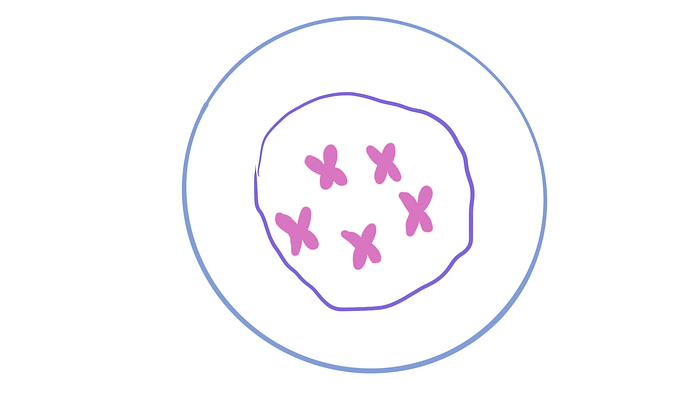
Then in late prophase, the nuclear envelope begins to disassemble and each chromosome is connected to a spindle fibre at its centromere. Spindle fibers are connected to the centrioles on each side. The centrioles are like the military establishments while the fibres are the commanders, guiding the soldiers. Although this part doesn’t align with the analogy, the chromosomes don’t actually move on their own at all and it’s the fibres doing all of the work!

Next comes metaphase where the chromosomes line up in the middle and the nucleus is fully disassembled. Here the mitotic spindle’s completed and the tubes connect centromeres to poles. This is where the army is starting to organize and their training becomes complete.
And the mitotic spindle is just the collection of the spindle fibres!
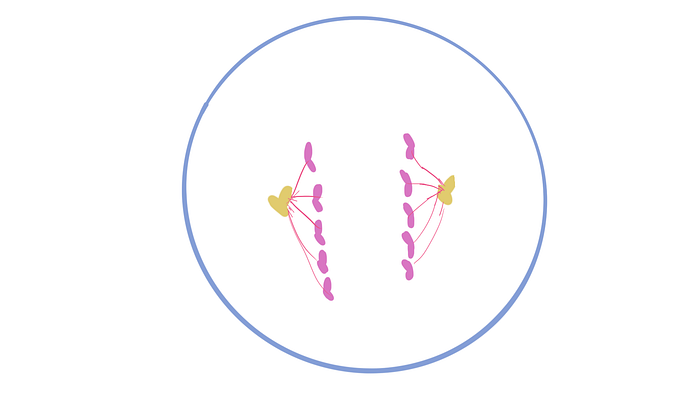
After that is anaphase; try remembering it with “a for away” because this is when the sister chromatids separate at centromere and they are now each a complete chromosome. Remember that the number of centromeres is what matters so there are now double the number of chromosomes! The separated chromosomes are pulled to opposite ends of the cell! This is when the trained soldiers are being sent to different battles.
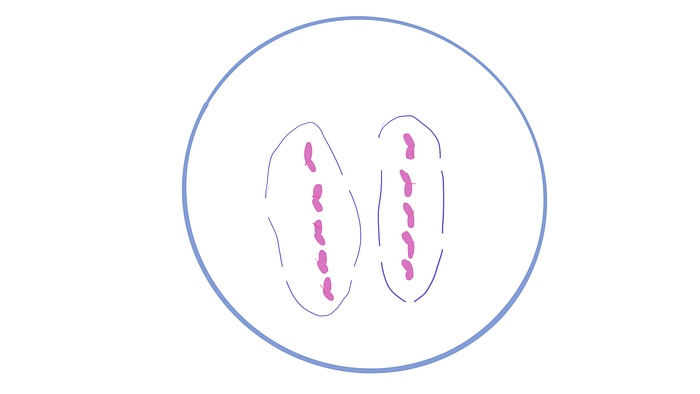
And now it’s time for telophase! This is when the chromosomes are at complete opposite ends of the cell and the centrioles begin to disappear. As well, the nuclear envelope starts developing. The soldiers are nearly ready for battle.
Finally, in cytokinesis, a process outside of mitosis but the step where the actually division of the cell happens, occurs! It’s probably the most interesting one!
Let’s see this in action!
Apical vs. Basal
This is a pretty simple concept but one that’s going to be pretty important later on. The apical side is the one facing the external environment while the basal side is the one that’s attached to an underlying surface or tissue.
Take a look at the diagram below to really solidify the idea!

What’s a Protein Complex?
It sounds simple enough but this is another important concept so let’s cover it quickly.
A complex is just a group of two or more polypeptide chains that come together to form a unit. Without getting into too much detail polypeptide chains are chains of amino acids. And amino acids are the building blocks of proteins.
Imagine you’re creating necklaces by threading beads onto a string. You then get a few of these necklaces tangled up together. Voila, you’ve got yourself a complex!
Asymmetric Cell Division
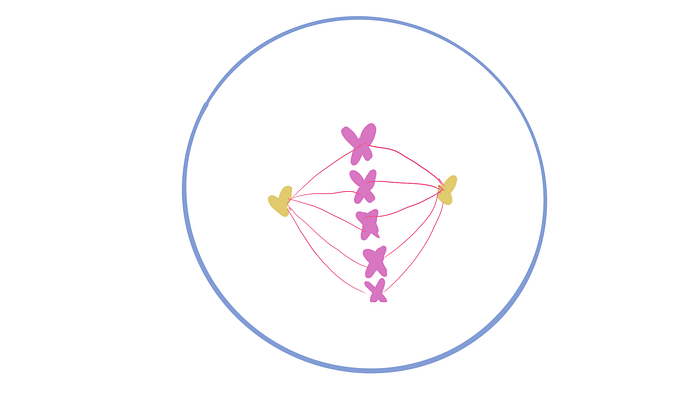
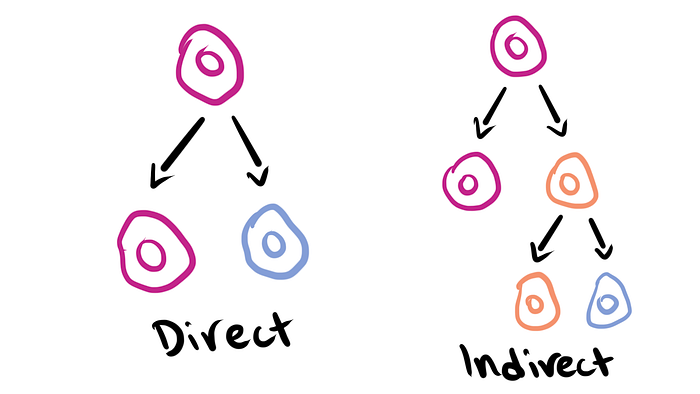
Now that the cell cycle’s out of the way, we can get to the exciting part! We’ve already learned the difference between symmetric and asymmetric cell division (or ACD for short) but there are actually two subtypes of ACD: direct and indirect.
Direct is when there are just two daughter cells: one that carries on the same as the mother and one that’s different. But indirect is when one takes after the mother cell while the other is an intermediary of sorts which then undergoes the cell cycle itself to produce more specific cell types. Look at the picture below to get a better idea of what that means!
A lot of what we know about ACD comes from fruit fly or Drosophila neuroblasts which are essentially the stem cells of the Drosophila central nervous system.
Let’s back up a second though; when do we need ACD and when do we need the symmetric version? Well, like I mentioned, stem cells are one of the main cells taking advantage of ACD. I won’t go into too much detail about what stem cells are (you can check out my video on them here) but let’s quickly go over their characteristics. To put it simply, they have to check off 2 boxes.
- They must be able to differentiate. That means they can turn into more specific cell types, like a hematopoietic stem cell or blood stem cell becoming a T cell.
- They must be able to self-renew. This is pretty self-explanatory: they can make identical copies of themselves.
In order to both make identical and different copies, stem cells utilize ACD.
Now the first thing we need to understand about ACD is what cell polarity is! This paper describes it as “an asymmetry in molecular composition or structure between two sides”. Cell polarity is essential to ACD but based on our current research, it’s a step that comes before it. In other words, ACD doesn’t cause cell polarity. Rather, the polarity is “decided” by the cell before we get into any of the ACD mechanisms!
And the last thing we should talk about before getting into the causes of ACD is mitotic spindle orientation. It’s exactly what it sounds like: the way the spindle is oriented. A while ago, we used to think that spindle orientation and the localization of some fate determinants (these are proteins or other molecules that can change the fate of the daughter cell) were dependant on each other. However, we now know that while both events are necessary for ACD, they happen independently of each other!
With all of that out of the way, what are the causes of ACD?
Intrinsic Factors
Numb
Numb is a signalling inhibitor which means it stops signals. In this case, Numb suppresses Notch signalling which we’ll learn about in a bit
It’s actually evenly spread all throughout the cytoplasm during interphase but during mitosis, it begins to localize, meaning it starts gathering in a specific place like you can see in the video on the left. Numb actually always localizes near one pole of the mitotic spindle.
In stem cells, the one sister cell that gets the higher levels of Numb suppresses signals and differentiates. The one with the lower level takes on the same stem cell fate as the mother cell.
Inscuteable
Remember how I mentioned the importance of orientation in ACD? Well, Inscuteable, or Insc for short, plays a vital role in reorienting the mitotic spindle!
Normally, in the absence of Insc the mitotic spindle is parallel to the surface the cells are attached to. But when Insc is there the spindle becomes oblique, or slightly tilted to one side.
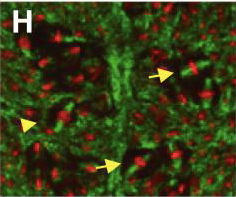
Take a look at how this works in mice cells. The mitotic spindles are in green and the DNA is in red. The yellow arrows are pointing to cells where the spindle is oblique.
And when Insc was overexpressed in mice, there was an increase in the ratio of oblique to non-oblique cells!
And they also found that Insc played a large role in creating intermediary daughter cells, the ones I mentioned that would go on to give rise to more specific cell types. These intermediary cells were most likely to occur from mother cells with oblique or vertical mitotic spindles.
You can think of the mitotic spindle as a knob and Insc as the force moving it. When the knob is perfectly horizontal, everything is “normal” but as it gets turned by the force (the Insc), things start to change and ACD takes place!
Now when the Insc helps the spindles orient fully vertically, it gathers or localizes in the apical side! But how does it know to do that? It’s not like Insc can actually think for itself… right? No, it cannot think for itself; there’s actually something much cooler going on.
Partner of Inscuteable (Pins) / LGN & Gαi
Pins really is the partner of Insc because it helps it do its job quite well! And LGN is just the equivalent in humans while Pins is for Drosophila.
Pins is responsible for stabilizing the mitotic spindle once it’s in position and maintaining the correct polarity and orientation. And Pins is also found at the apical side of the cell along with Insc.
Based on my research, Gαi plays a very similar role to Pins/LGN in ACD. It’s also found alongside Insc in the apical side.
Mud/NuMA
These are microtubule binding proteins, with microtubules being like the little arms that pull the chromatins during anaphase; Mud is found in Drosophila and NuMA is the mammalian version. So what they do is help attach the mitotic spindle to a part of the cell and help with maintaing the orientation of it!
They’re a part of the Pins and Gαi protein complex and they’re pretty cool. In epithelial cells (those are the ones that line cavities and form thin tissues) when there’s no Insc, the mitotic spindle orients itself horizontally and the Mud/NuMA help create microtubule attachments on each horizontal end.
But when Insc is present, things are different. Insc gathers at the apical side and so does the Pins, Gαi, and Mud/NuMA protein complex. So now the mitotic spindle is not only oriented vertically because of the Insc but there are also microtubules that attach is that way.The Par Proteins
They’re like one of those power families that are all super successful and get things done! In this case, they were found to be responsible for the unequal sharing of the mitotic spindle in C. elegans.
But this isn’t just in C. elegans; we’ve found that the Par proteins: Par-1, Par-2, Par-3, Par-4, Par-5, and Par-6 are somehow involved in nearly anything related to cell polarity.
The most important Par proteins that form a complex vital to ACD are Par-3, atypical protein kinase C or aPKC, and Par-6.
With the Par complex though, the location doesn’t change with regards to whether Insc is present or not. But since the complex is always located at the apical side, it doesn’t do much when Insc isn’t present and the mitotic spindle is horizontal.
It’s only when Insc is present and the spindle is vertical that the Par complex gets to shine because it gets attached to the spindle.
A Focus on aPKC
All of the Par proteins are pretty cool but let’s focus on aPKC right now because it can do something really cool. Recall how Numb was a fate determinant that was unevenly split between the two sister cells. Well how does that segregation happen?
When a mitotic kinase called Aurora-A becomes active, it phosphorylates the Par-6 part of the Par protein complex which in turn makes the aPKC phosphorylate the Numb. Put differently, aPKC is inhibited by Par-6 until Aurora-A phosphorylates Par-6.
This neutralizes the charges and the Numb protein basically falls from the apical side to the basal side of the cell. Now instead of being evenly spread around the perimeter of the cell, it’s mostly concentrated in the basal side!
Think of it like this. You and your friends have made a circle together and are holding hands. But suddenly someone pushes all of the people on the top side and they fall over to the bottom. Now there’s a concentration of people at the bottom. A bit of a strange analogy but it works!
Back to the science, what’s phosphorylation or a kinase? Phosphorylation is just when a phosphate group is introduced into a molecule and a kinase is an enzyme, a catalyzer.
Notch Signalling
Notch signalling is unequal in the two sister cells because of the different amounts of Numb in each of them.
Notch plays a critical role in differentiation in both humans and in Drosophila. In fact, one study found that the inactivation of Notch signalling resulted in a lack of ACD where both daughter cells divided symmetrically! Even more interestingly, the times at which Notch was inactivated mattered. When this was done early on, the Numb divided symmetrically. But when it was inactivated later, just made the asymmetric segregation of Numb incomplete.
Extrinsic Factors
Niche
While there were a ton of intrinsic, the one major extrinsic factor that you’ll hear most about with regards to ACD is stem cell niches. And thankfully, it’s a fairly simple concept! A stem cell niche is a microenvironment where stem cells are and it’s where the stem cells are provided with a lot of critical signals and factors so they remain stem cells.
These niches have a limited range as to how far their signals go, ensuring that only the cells that are close by receive the signals, while those farther away differentiate and lose their stem cell potency.
So when a stem cell divides asymmetrically, one daughter cell would be inside the niche while the other wouldn’t be. The one inside would also follow the stem cell path while the one outside would differentiate.
Tying It All Together
Now bringing it all together, I want to hint back at the title of the article which if you don’t remember is “Twins Separated At Birth: The Story of Asymmetric Cell Division”. It’s a pretty cool analogy so buckle your seatbelts.
There are two identical twins who are separated at birth. One of them stays with their genetic parents and so ends up very similar to them, not just genetically but also in terms of habits and behaviours. The other one goes to live with a super different family and so differentiates and ends up pretty different from the other twin even if they came from the same parents.

That’s the story of ACD or at least the one involving stem cell niches; if you’re ever forgetting how niches work just remember the Cheeto and broccoli twins!
Want More?
And finally the above article was pretty awesome too but beware that it goes pretty deep!
Hey there 👋 Parmin here; I’m a 15 y/o student studying stem cells at The Knowledge Society 🧪 Everyday, I aspire to uncover the secrets of biology and learn something new! Make sure to follow me on Medium to hear about every new article I post, connect with me on LinkedIn, or contact me at parminsedigh@gmail.com! Also subscribe to my monthly newsletter to learn about every cool, new thing I’m working on ✍️
References
Feel free to check out all of my references right here.
